
Крупные человеческие сообщества исторически организованы как государства и цивилизации — являются сложными социальными системами, которые отличают кооперативные макроэффекты, отсутствующие на индивидуальном уровне. Потому, например, Робинзон Крузо на острове не представлял собой сложной социальной системы.
В рамках авторского научного поиска основополагающим тезисом выдвигается представление, что человеческое сообщество, будучи самостоятельной целостной формой жизни, наследует все эволюционные законы развития, которые присущи и биологической жизни как таковой. Более того, такое представление социальных сообществ позволяет усмотреть дальнейшее видоизменение процесса эволюции жизни. За биологической изменчивостью с неизбежностью следует изменчивость социализирующая и затем — нравственная изменчивость. Наряду с антропогенезом и социогенезом, таким образом, следует видеть и предвосхищать следующую стадию эволюции жизни, а именно — сологенез.
Этой новаторской обществоведческой проблематике посвящена монография проф. В.Э.Багдасаряна и С.С.Сулакшина «Сложная социальная система в витальном подходе» (Москва: Научный эксперт, 2013).
Приводим ниже фрагмент 2.2 «Расово-генетическая компонента цивилизационогенеза» из раздела «Витальные факторы, воздействующие на сложную социальную систему», посвященный расово-биологическому началу и его влиянию на развитие цивилизационных общностей.
Первая часть публикации доступна здесь.
ФАКТОР ГЕНЕТИЧЕСКОГО СКРЕЩИВАНИЯ
Долгое время в биологии доминировало представление о нескрещиваемости видов. Гибриды, считалось, не способны к производству потомства. Однако еще К. Линней допускал возможность гибридогенного видообразования[87]. Голландский генетик Я.Лотси пошел дальше, рассматривая гибридизацию как основу эволюционного процесса[88]. Подобно тому, как индивид имеет двух родителей, рассуждал он, так и каждый вид генетически восходит к двум предковым видам. В СССР сторонником этого подхода выступал филогенетик М. Г. Попов[89]. Согласно ему, эволюционное развитие следовало бы представлять не в виде традиционного дерева, а в форме сети.
Каждый из таксонов в ней может взаимодействовать с другими, формируя новый биологический вид. Экспериментально возможность гибридогенного образования была подтверждена в трудах селекционеров Г. Д. Карпеченко, И. В. Мичурина, А.Мютцинга, Д. Л. Стеббинса и др.[90] Развитие этого направления теории эволюции привело к констатации существования в природе наряду с описанным Ч.Дарвином дивергенционным, конвергенционного пути создания новых видов. Было установлено, что видообразование может осуществляться не только в направлении разделения видовых линий, но и их синтеза[91].
Все эти положения применимы и к вопросу об антропогенезе. Современные исследования генома человека четко фиксируют многочисленные факты межрасовых и межэтнических скрещиваний. На их основе формировались генетически новые человеческие популяции. На биолого-генетической основе выстраивалось далее здание специфичного социального бытия. Возникали этносы. Каждый из них соотносился, как правило, с особым вариантом геномных комбинаций. Культурная традиция находилась в эпоху традиционного общества в интегральном единстве с соответствующим генотипом. Локализованность бытия этносов в этот период определяла генетическую эндогенность, а соответственно, и устойчивость генофонда. Брачная замкнутость нарушалась только в периоды войн и переселений народов. Такого рода миграции вызывали соответствующие волны ускорения этногенеза.
Процесс глобализации породил новую реальность в геномных изменениях человека. Расширение международных коммуникаций, с одной стороны, объективно разрушает этнические геномы. Брачная замкнутость этносов, хотя и остается значительной в отдельных регионах, в целом в мире уходит в прошлое. С другой стороны, мировые миграции носят теперь преимущественно индивидуальный характер относительно существовавшего ранее формата целенаправленного переселения народов. Отсюда прежняя модель этногенеза, когда скрещивание двух этносов вело к формированию нового генома, более не действует. Генный обмен приобретает открытый в планетарном масштабе характер. В соответствии с этими тенденциями футурологическая перспектива видится в генетической конвергенции человечества. Геномные различия этносов будут со временем все более ослабевать. Конвергенция человечества на биологическом уровне идет одновременно с процессом социальной конвергенции цивилизаций.
Применяя к социальным системам модель биологического формообразования, можно предположить, что новые этносы способны возникать в процессе как популяционной дивергенции, так и конвергенции. Дивергентный процесс связан с адаптацией отдельных групп единого прежде этноса к специфическим средовым условиям бытия. Об этих адаптационных механизмах говорилось в предшествующем разделе.
В данном случае более интересен феномен этнических синтезов. На первый план здесь уже выходит вторая основополагающая характеристика популяций — специфичность их генетической природы. Эта специфика применительно к возникновению этносов достигается посредством вариаций межэтнического скрещивания. Факт такого синтеза обнаруживается на ранней фазе истории большинства народов. Римская история открывается преданием о похищении первыми римлянами сабинянок, ставших их женами. Этногенез европейских народов выстраивался в синтезе германского, романского и кельтского компонентов. Русская летописная традиция берет в качестве отправной точки призвание новгородскими славянами варягов. Приведшие к формированию евразийской империи завоевательные походы Чингисхана начались с этнического монголо-тюркского синтеза, отразившегося в появлении этнонима татаро-монголы. Фактор этнического смешения раскрывается при соответствующем анализе как универсальный опыт этногенеза. Это прямо опровергает расистские мифы о «расовой чистоте». Этносы, как и новые геномы, исторически возникали в результате популяционных смешений. Генетически чистых, несмешиваемых общностей никогда не существовало.
Геномная природа любого народа по меньшей мере дуальна. Конечно, в ней может быть доминирующий генный компонент. Но само образование соответствующей этнической группы определялось не этой доминантой, а ее смешением (табл. 2.2.1)[92].+
Таблица 2.2.1. Фактор этнического смешения в этногенезе
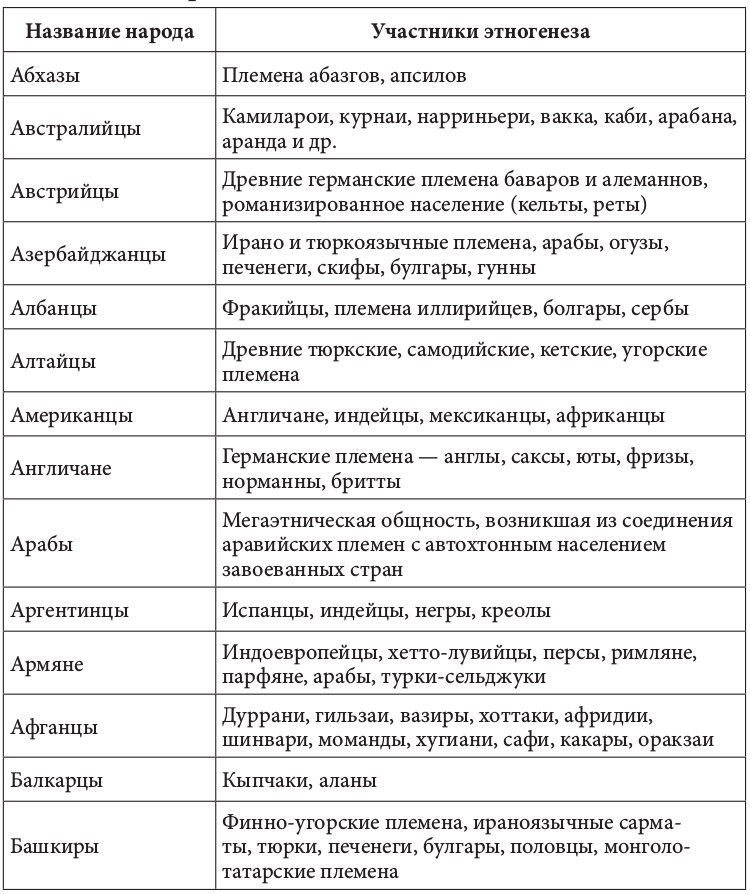
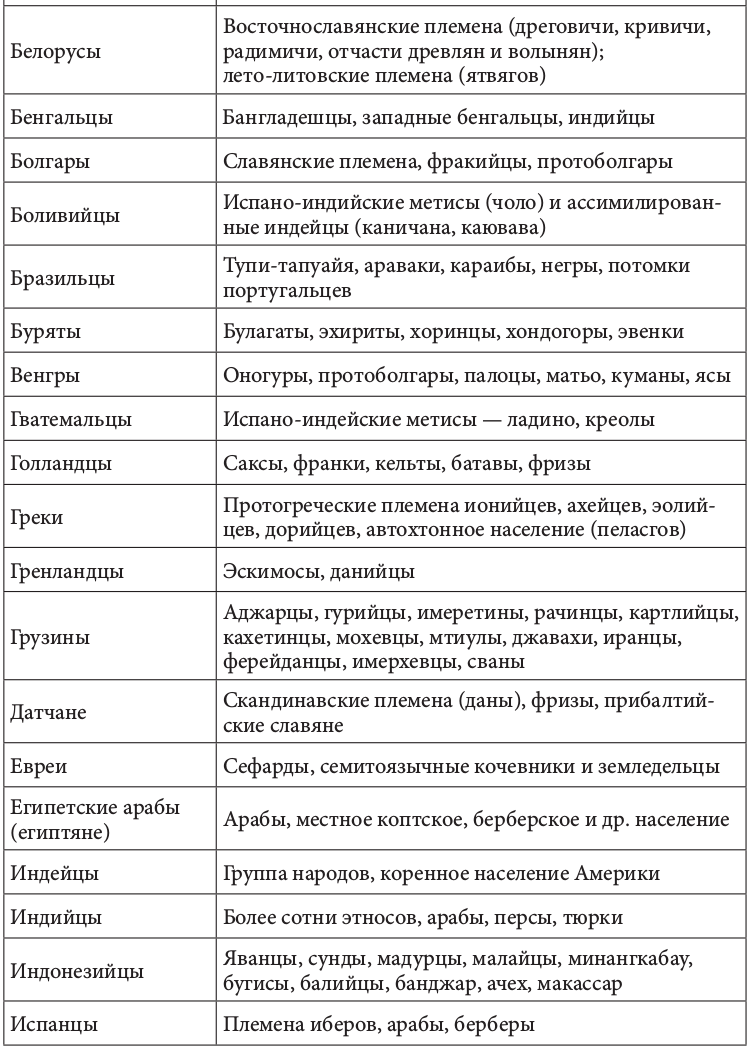
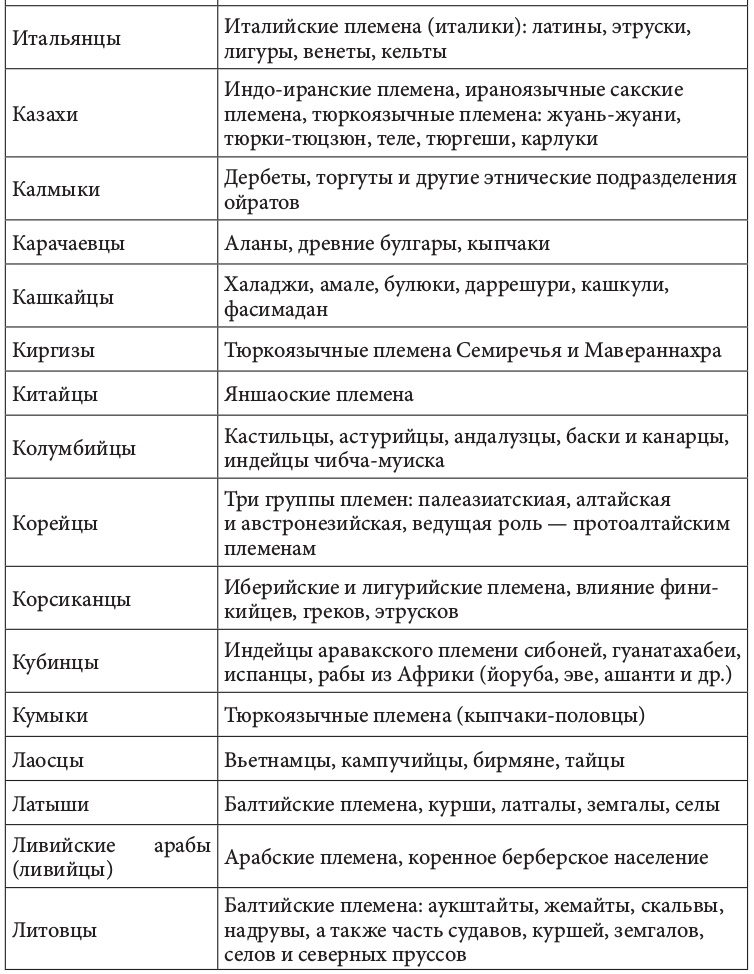
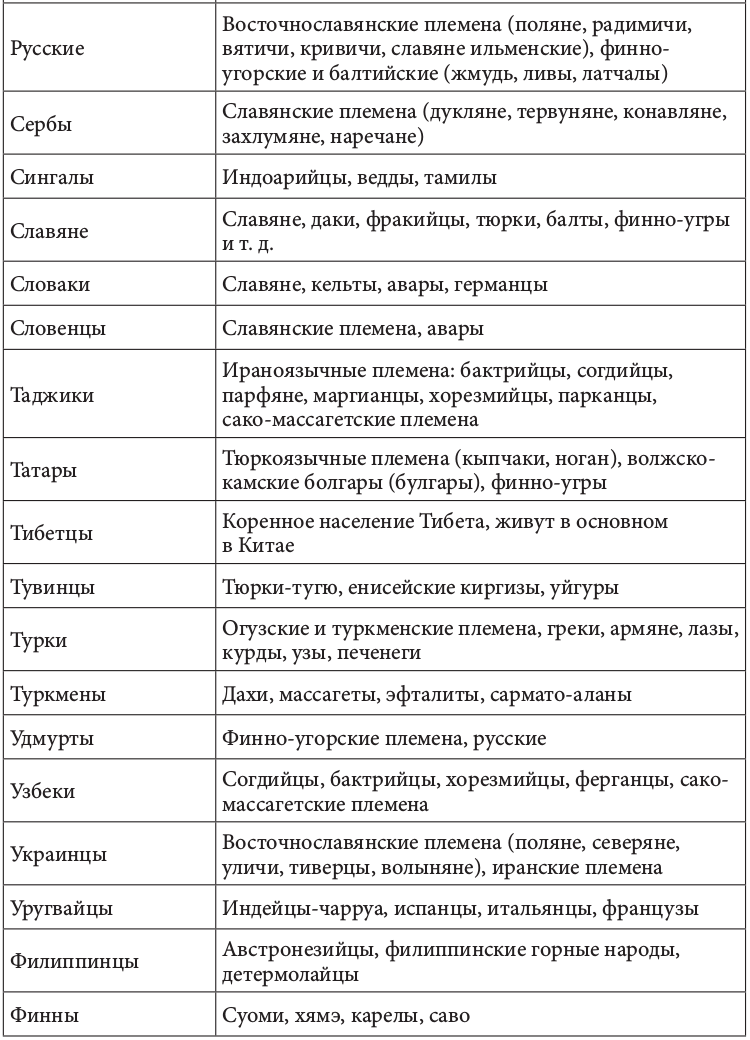
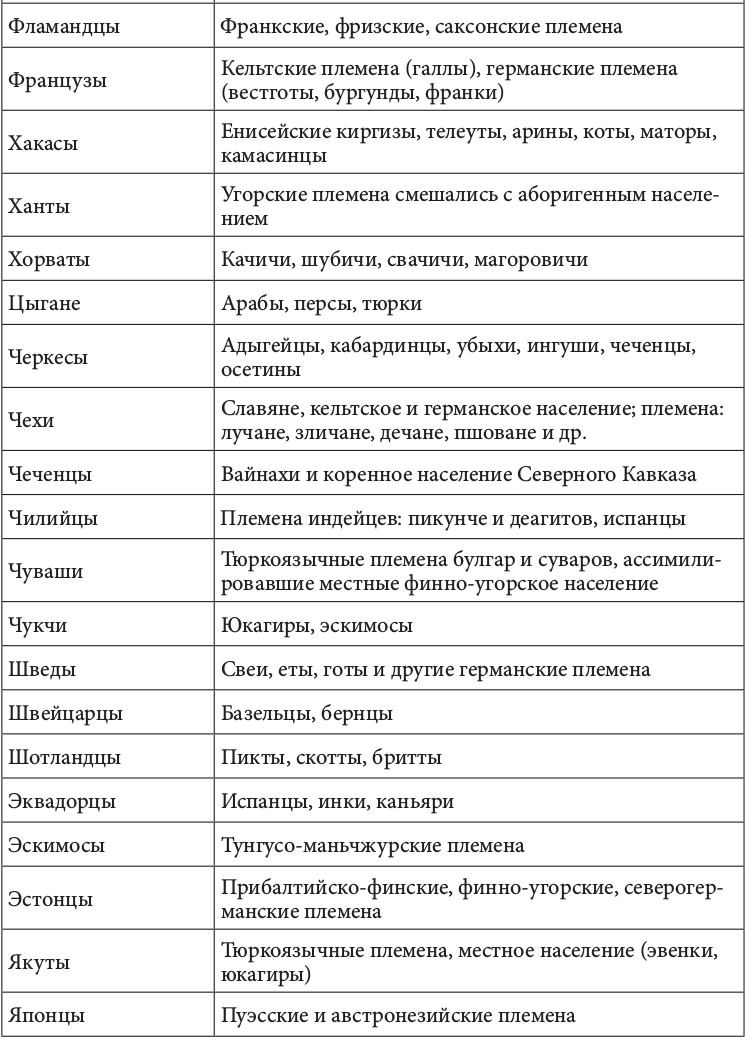
Движущей силой генных трансформаций исторически выступали расовые и этнические миграции. Прослеживается прямая связь между ними и начальной фазой цивилизационогенеза. Цивилизация в Европе начала складываться после перемещения туда земледельцев из Передней Азии, зоны «плодородного полумесяца». Они совокупно составляли не более 10% европейского населения. Но их приток вызвал утверждение новой земледельческой парадигмы хозяйствования. И именно эта волна миграции, как утверждают генетики, определила формирование основных черт европейского генофонда. Генетическими исследованиями установлено также, что формирование ранней средиземноморской культуры соотносилось с широким распространением по всему Средиземноморью финикийского генотипа. Индоарийские миграции начала II тыс. до н. э. вызвали появление ведической индийской и авестийской иранской цивилизации[93]. История становления китайской цивилизации представляла череду вторжений различных по этнической принадлежности кочевых племен. Генетически прослеживается наличие в цивилизационогенезе Китая и европеоидного компонента. Расово смешанным европеоидно-монголоидным считают историки состав населения древнейших китайских культур Яншао и Шан-Инь[94].
Геном древнего эллина сформировался в результате смешения трех этнических компонентов — ионийского, ахейского и дорийского. Ахейцы и дорийцы выступили первоначально по отношению к автохтонному населению в качестве завоевателей. Ахейцами были, в частности, афиняне, дорийцами — спартанцы. Неассимилируемость спартанцев с автохтонами — илотами была скорее исключением в рамках общегреческого процесса генетического смешения[95]. На Западе индоевропейская миграция начала II тыс. до н. э. привела к формированию хеттской, урартской и эллинской цивилизации. Походы Александра Македонского имели для Ближнего Востока принципиальное значение как фактор не только культурной, но и масштабной генетической трансформации.
Волны завоевательных миграций привели к исчезновению генотипа населения древних цивилизаций — древнеегипетского и древнешумерского. Так, Египет был последовательно завоеван гиксосами, ассирийцами, персами, греками-македонцами, римлянами, арабами, турками и англичанами. Понятно, что исходное геномное основание древнеегипетского цивилизационогенеза оказалось принципиально изменено. Генетики полагают, что наиболее близки к древним египтянам современные копты, составляющие ныне не более 10% населения Арабской республики[96].
С «исхода» народа Моисея из Египта в Палестину традиционно ведется отсчет формирования еврейского цивилизационного анклава. В период «рассеяния» религиозные группы евреев придерживались жесткой традиции брачной замкнутости. Это позволило им избежать естественной ассимиляции. Однако генетический обмен все же осуществлялся. Как следствие, возникшие субэтнические группы, евреи-ашкенази и евреи-сефарды имеют заметные фенотипические различия. Другие иудейские этнические локалитеты: горские евреи, бухарские евреи, феллахи, таты, караимы, крымчаки, — и вовсе принадлежат к иным расовым типам. Соответственно, несмотря на общность религии, под единым маркером иудаизма сложились существенно варьирующие культурные модели. Неслучайно современная эпоха, наступившая после разрушения Иерусалимского храма, характеризуется в иудаике как «эпоха смешения»[97].
Древние римляне утверждали, что их прямыми предками были выходцы из разрушенной ахейцами Трои. Идея реванша над греками поддерживалась ими весь период античности и была передана в средние века западноевропейским католикам. Ранняя фаза римского цивилизационогенеза определялась многообразием этногенетических компонентов, основные из которых были представлены греческими колонистами, италийцами и этрусками. Этномиграционное содержание периода перехода от античности к Средневековью в Европе не нуждается в комментариях. За этим временем в исторической литературе прочно закрепилось понятие «великого переселения народов». Инерция глобальных миграционных сдвигов действовала около пяти столетий. Итогом этих процессов стало принципиальное переформатирование генетической и цивилизационной карты Европы и значительной части Азии. Римляне как этногенетический тип исчезли.
Одним из итогов «великого переселения народов» стало формирование и славянской этничности[98].
Исторически становление исламской цивилизации было связано с арабской миграционной экспансией. Сегодня территория расселения арабов простирается от Марокко на западе до Ирака на востоке. Понятно, что в каждом из географических локалитетов происходило их генетическое смешение с разнородными группами автохтонов. Генетически значимым было участие арабов в этногенезе испанцев, португальцев, итальянцев, сардинцев. Другой миграционной волной, сыгравшей принципиальное значение в развитии цивилизации ислама, является западная турецкая экспансия. На развалинах Византии была воздвигнута Османская империя. Не арабы, а турки выступили на определенном этапе в качестве новой цивилизационнообразующей силы. Сегодня под единым маркером исламской цивилизации существуют-де-факто три центра силы, четко соотносимые с тремя компонентами этничности: арабским — саудитским, тюркским — турецким и индоарийским — иранским.
Современное японское общество, казалось бы, этнически гомогенно. Однако созданию первого общеяпонского государства Ямато предшествовал процесс генетического смешения переселившихся с Корейского полуострова пуесских монголоидных племен с коренным айноидным и австронезийским населением. Переселенцы принесли с собой поливное земледелие, определив формирование жизненного уклада будущей японской цивилизации. Процесс ассимиляции продолжался до XIX в. Окончательно японский генотип сложился только на фазе цивилизационного подъема, связанного с революцией Мэйдзи[99].
Генезис русской цивилизации соотносится с распространением славянского генотипа в VIII—IX вв. в направлении с юго-запада на северо-восток. Фиксируется также активное смешение его с финно-угорским и балтийским компонентом. Вместе с тем генетическими исследованиями не подтверждается распространенное прежде представление о значительной монголоизации русских под влиянием ордынского ига. Суммарная частота восточно-евразийских гаплогрупп в русских популяциях не превышает 2%. Выдвинутый в свое время на Западе миф о монголоизации решал вполне определенные политические задачи. Необходимо было дезавуировать в глазах европейцев российскую цивилизацию, связав ее с «азиатчиной» и «варварством»[100]. К тектоническим цивилизационным сдвигам привела, очевидно, начавшаяся с эпохи Великих географических открытий колониальная экспансия Запада. Но в контексте рассматриваемой проблематики важно ее соотнесение с геномными трансформациями. Генные изменения вносятся еще миграциями периода крестовых походов.
У современных ливанцев генетики четко фиксируют европейские генотипические примеси. После открытия Америки с конца XV в. начался незавершенный до сих пор процесс формирования на американском континенте новых геномов. В Южной Америке, где между колонистами испанцами, португальцами и автохтонным индейским населениям не было жесткой брачной некомплиментарности, этногенез осуществлялся быстрее[101].
Метисы сегодня являются преобладающим расовым типом на южноамериканском континенте. В Венесуэле их удельный вес достигает 92% общей численности населения. Новый генотип формируется буквально на глазах. Наряду с исходными европейским и индейским компонентами определенную роль здесь сыграли негритянский и индийский генетические элементы. Мулаты по сей день численно преобладают в Гвиане, а индийцы — в Гайане и Суринаме. Южноамериканский опыт иллюстрирует в данном случае общую модель этногенеза как проявления этногенетического смешения. И вот уже все отчетливее проявляется факт существования особой цивилизации Латинской Америки. Новое геномное формирование совпадает, таким образом, с процессом цивилизационогенеза. Характерно, что именно Венесуэла, наиболее метисизированная страна континента, претендует сегодня на роль цивилизационного лидера (рис. 2.2.10).
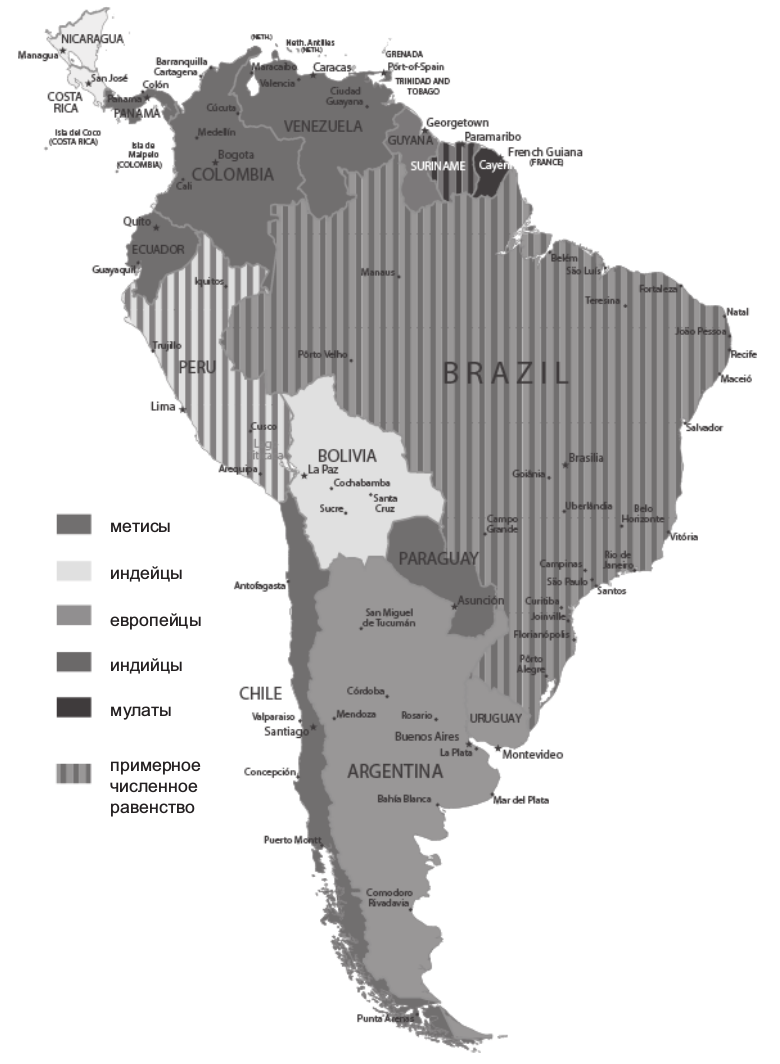
Рис. 2.2.10. Карта расового доминирования в странах Южной Америки
В США, как известно, давно на уровне идеологического выбора был выдвинут концепт этнического плавильного котла. Де-факто вопреки этой установке долгое время действовали принципы расовой сегрегации. Этнический котел ограничивался главным образом потомками выходцев из Европы. Однако со временем внутрирасовые границы брачности все более размываются. Символом нового этапа этнообразования является фигура президента США Барака Обамы. Генеалогия его по линии отца — это кенийский народ луо, по матери — англичане, шотландцы, ирландцы, немцы и индейцы чероки. Геном Б.Обамы не поддается локализующей этнической идентификации[102].
Глобализация и многовекторность миграционных потоков позволяет применительно к сегодняшнему дню говорить о перспективе геномной конвергенции человечества. Уже сейчас в мире количество мигрантов превышает 175 млн чел. Но геномная конвергенция произойдет не завтра. Пять из шести жителей планеты проживают еще в том месте, где родились[103]. Первым в фазу сущностных генетических трансформаций вступит западный мир. Доля мигрантов в нем по отношению к общей численности населения находится в диапазоне от 9,1% в странах Европы до 20,5% в Австралии. Для сравнения, в Китае их удельный вес на сегодня составляет лишь 0,05% (рис. 2.2.11)[104].
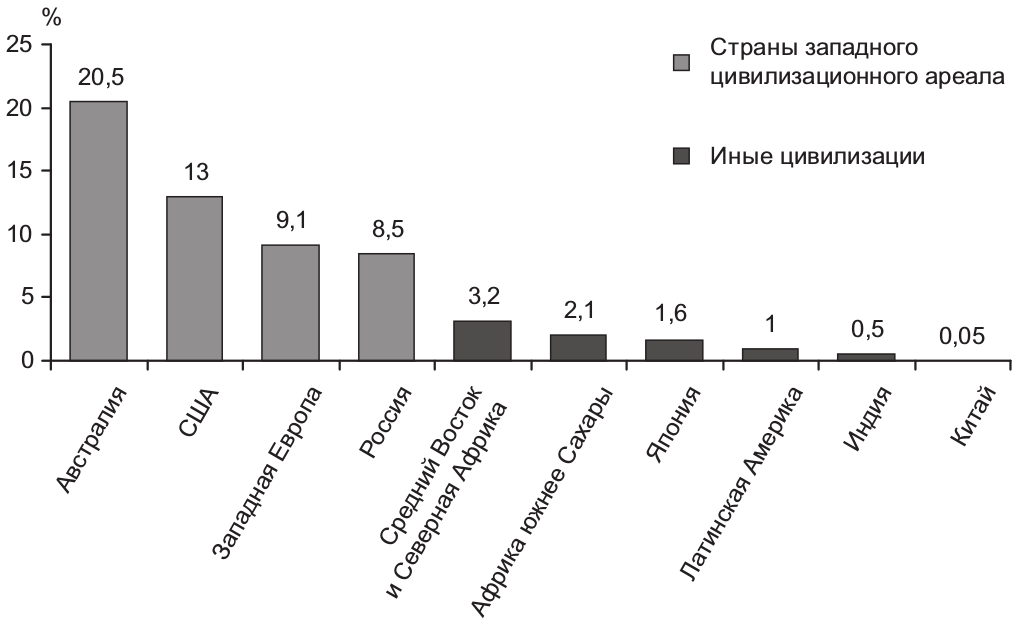
Рис. 2.2.11. Доля мигрантов в численности населения по ряду цивилизаций и стран мира
Миграционное давление на Запад, имея в виду усугубляющиеся цивилизационно-страновые диспаритеты в качестве жизни, будет в дальнейшем только возрастать. Исходя из полученного вывода о зависимости цивилизационогенеза от популяционно-генетических изменений, следует предположить, что грядущая геномная трансформация приведет к существенной трансформации западной цивилизации. Формируется генетически новый тип человека, для которого прежний набор ценностей Запада может оказаться неприемлемым.
О РАСОВОМ РЕДУКЦИОНИЗМЕ
Влияние генно-популяционного уровня бытия жизни на развитие цивилизационных систем, таким образом, существует. Однако действие его ограниченно. У А. С. Пушкина, как известно, была по линии деда Ибрагима Ганнибала примесь африканской крови. М. Ю. Лермонтов имел по отцу шотландские корни, а по матери — татарские. Матерью В. А. Жуковского была пленная турчанка. Род Толстых вел свое происхождение от немецкого иммигранта XIV в. в Чернигов «мужа честна Индриса», выехав шего «из Цесарския земли». В XVIII в. переехал на службу в Россию из Германии предок А. А. Блока. Отец В. А. Даля был датчанин, мать — француженка. Род Сумароковых ведет свое происхождение по одной версии из Швеции, по другой — из Литвы.
Г. А. Державин сам воспел в стихах свое татарское происхождение. Н. М. Карамзин гордился производностью своей фамилии от некоего Кара-Мурзы. К татарской знати принадлежали предки И. А. Тургенева и И. А. Куприна. Польских предков имели великие русские писатели А. С. Грибоедов, Н. В. Гоголь, Ф. М. Достоевский, Н. А. Некрасов, И. А. Бунин. Но это никоим образом не помешал и Гоголю, и Достоевскому резко высказываться против Польши. При генной идентификации авторов русская классическая литература вообще перестала бы быть русской. То же относится к другим сферам бытия. Такая же генетическая гетерогенность обнаруживается в отношении других национальных культур. Но все вышеназванные литераторы составляют лицо русской творческой идентичности. Творчество их внесло сущностный вклад в развитие национальной саморефлексии. Значит более значима не связь с этническими генами, а ценностный выбор человека. Это подтверждает выдвинутый выше тезис о возрастающей приоритетности духовного уровня бытия жизни над биологическим.
Неправомочность биолого-популяционного детерминизма следует и из данных самой генетики. Рисунок 2.2.12 [105] иллюстрирует значимость различных уровней принадлежности человека в его геномной структуре.
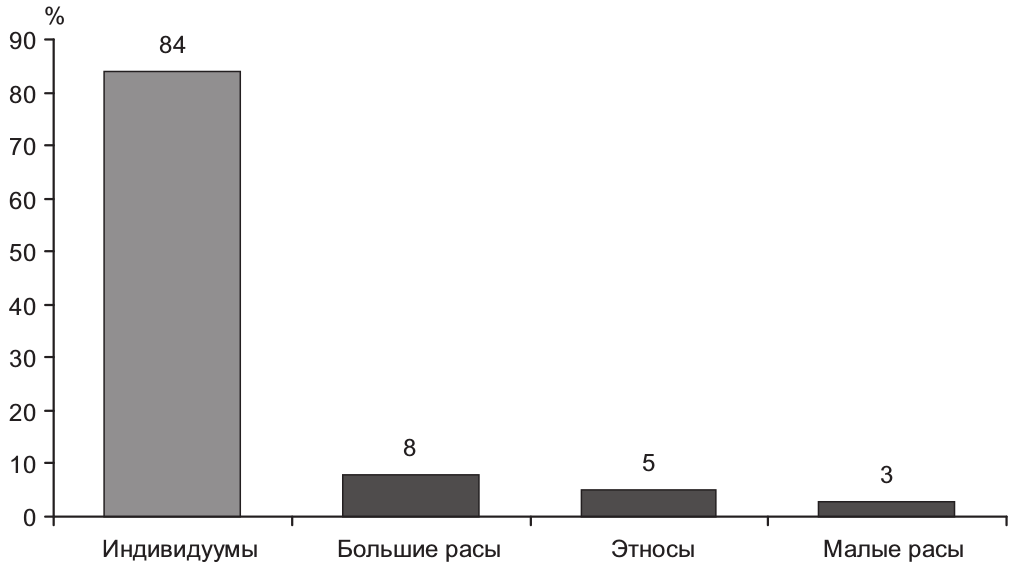
Рис. 2.2.12. Доля генетического разнообразия в зависимости от расово-этнической принадлежности человека
Нельзя сказать, что расовая или этническая составляющая не играет никакой роли. Но эта роль не превышает совокупно 16%. Индивидуумные различия людей оказываются в геноме человека гораздо более значимы. Следовательно, цивилизационогенез расово не детерминирован. Расистский редукционизм, так же как и географический, научно не подтверждается.
Используя параметр генетического расстояния, современная генетика позволяет количественно (без расистских мифологем) оценить степень различия рас в общей мегаэволюции мира[106].
Она позволяет также ответить на вопрос, с каким из типов биологических топосов соотносятся сложные социальные системы, такие как цивилизации. Величина расстояния для больших человеческих рас составляет 0,03%. Много это или мало? Для сравнения: межвидовое расстояние составляет от 0,5% и выше. Диапазон генетического различия подвидов от 0,17 до 0,22%. Таким образом, ни видами, ни подвидами расы не являются. Зато показатель генетического расстояния в 0,03% соответствует в животном мире уровню местных популяций. Следовательно, популяция есть на языке биологического бытия категория, наиболее адекватная (аналогичная) для цивилизационных систем[107]. Беллетристически говоря, действительно — «все люди братья».
Возможность применения популяционного подхода к пониманию биологического уровня бытия сложных социальных систем, таким образом, подтверждается. Вместе с тем выявляется принципиальная недостаточность только биологической апелляции в объяснительных моделях сложной социальной системы. Это приводит к задаче проследить факторную связь биологического и духовного уровней существования человека, предположив доминирующее влияние последнего в процессе цивилизационогенеза.
ПРИМЕЧАНИЯ
[87] Линней К. Философия ботаники. М., 1989. С. 352.
[88] Lotsy J. P. Evolution by means of hybridization. Hague, 1916.
[89] Попов М. Г. Основы флорогенетики. М., 1963; Его же. Географо-морфологический метод систематики и гибридизационные процессы в природе // Труды прикладной ботаники, генетики и селекции. 1927. Т. 17. No 1. С. 221–290; Его же. Гибридизационные явления в природе и значение их для эволюции // Дневник Всесоюзного съезда ботаников. Л., 1928. С. 92–93.
[90] Карпеченко Г. Д. Теория отдаленной гибридизации // Теоретические основы селекции растений. М.- Л., 1935. Т. 1. С. 293–354; Мичурин И. В. Принципы и методы работы // И. В. Мичурин. Соч. Т. 1. М.-Л., 1939; Мюнтцинг А. Некоторые фазы эволюции тритикале // Проблемы экспериментальной биологии. М., 1977. С. 86–99; Стебиннс Дж. Л., Айала Ф. Х. Эволюция дарвинизма // В мире науки. 1985. No 9. С. 38–50.
[91] Воронцов Н. Н. Развитие эволюционных идей в биологии. М., 1999. С. 184–185; Грант В. Эволюционный процесс. Критический обзор эволюционной теории. М.: Мир, 1991.
[92] Народы мира: историко-этнографический справочник / Гл. ред. Ю. В. Бромлей. М., 1988.
[93] Бонгард-Левин Г.М., Грантовский Э. А. От Скифии до Индии. Древние арии: мифы и история. М.: Мысль, 1974.
[94] Васильев Л. С. Проблемы генезиса китайской цивилизации. М. 1976; Гумилев Л. Н. История Хунну; Грумм-Гржимайло Г. Е. Западная Монголия и Урянхайский край. Т. II. Л., 1926.
[95] Хэммонд Н.Дж. Пелопоннес (Стремление дорийских государств к господству, около 750–650 гг. до н. э.). // Кембриджская история древнего мира. Т. III. Ч. 3: Расширение греческого мира. М., 2007; Шмидт Р. В. Античное предание о дорийском переселении // Вестник древней истории. 1938. No 2.
[96] Брестед Д., Тураев Б. История Древнего Египта. Мн., 2003; Зеленев Е. И. Египет. Средние века. Новое время. СПб., 1999; Семенова Л. А. Салах ад-Дин и мамлюки в Египте. М., Наука, 1966; Коротаев А. В. Долгосрочная политико-демографическая динамика Египта: Циклы и тенденции. М., 2006.
[97] Немировский А.А. У истоков древнееврейского этногенеза. Ветхозаветное предание о патриархах и этнополитическая история Ближнего Востока. М., 2001.
[98] Алексеева Т. И. Этногенез восточных славян по данным антропологии. М., 1973.
[99] Киддер Дж. Э. Япония до буддизма: Острова, заселённые богами. М., 2003; Воробьев М. В. Япония в III–VII веках: этнос, общество, культура и окружающий мир. М., 1980; Суровень Д. А. Возникновение раннерабовладельческого государства в Японии (I век до н. э. — III век н. э.) // Проблемы истории, филологии, культуры. Магнитогорск, 1995. Вып. 2. С.150–175.
[100] Балановская Е.В., Балановский О. П. Русский генофонд на Русской равнине. М., 2007; Балановская Е.В., Балановский О. П. Генетические следы исторических и доисторических миграций: континенты, регионы, народы // Вестник ВОГиС. 2009. Т. 13. No 2. С. 401–409.
[101] Шемякин Я. Г. Об особенностях цивилизационного процесса в Новом Свете в эпоху конкисты // Цивилизации. Вып. 3. М., 1995; 500-летие открытия Нового Света: Исторические судьбы Латинской Америки. М., 1992; Верлинден Ч., Матис Г. Покорители Америки: Колумб, Кортес. Ростов н/Д, 1997; Кинтеро Лопес Р. Встреча цивилизаций или геноцид? // Общественные науки и современность. 1992, No 5; Диас дель Кастильо Б. Правдивая история завоевания Новой Испании. М., 2000.
[102] Мендел Д. Барак Обама. СПб., 2008; Нир Б. Барак Обама для начинающих. Все о новом президенте США. М., 2009.
[103] Юдина Т. Н. Социология миграции. М., 2006. С. 89.
[104] Кузык Б.Н., Яковец Ю. В. Становление интегрального экономического строя — глобальная трансформация XXI века. М., 2008. С. 72–73; World Development Indicators. Washington, 2007; World Development Indicators. Washington, 2008. P. 372–374.
[105] Биология. В 2 кн. / Под ред. В. Н. Ярыгина. М., 2007. Кн. 2. С. 155.
[106] Nei M. Mathematical models of speciation and genetic distance // Population Genetic and Ecology / еds S. Karlin, Nero E. N.Y., 1976; Животовский Л. А. Популяционная биометрия. М., 1991; Дерябин В. Е. Многомерная биометрия для антропологов. М., 1983.
[107] Биология. В 2 кн. / Под ред. В. Н. Ярыгина. М., 2007. Кн. 2. С. 156.


